Dr.
Gerald S. Hecht
Associate
Professor of Psychology
College
of Sciences
webmaster@psiwebsubr.org
PSYC 381 - Sensation
& Perception Exam 2 Study Guide
Vision
Vision is
the most complex of the senses, culminating in the high resolution,
color images of the higher primates. The pathway must handle such
demands as transduction of light into coherent and synchronous neural
impulses, binocular and more distant depth perception, motion in the
visual field or of the observer or both--in as near to real time as
possible...all in a wide range of focal lengths and light levels. The
receptor is constrained by the physical laws of optics. Motion not only
must be perceived but also tracked by coordinated eye, head, and body
movements as well as lens-iris accomodation. These paths ultimately
lead
into the realms of memory and mentation, where we recognize and
understand what is being seen...the so-called "mind's eye" which can be
tapped by dreams and hallucinations as well.
Structure
of the Eye
Proper
size and shape are extremely important for eye function.
- The
foremost layer of the eye is a
thin layer of conjunctiva that is
actually transparent
epidermis that reflects over the exposed surface of the eye from the
lining of the lids. Being epidermis, it has no blood supply, true of
structures generally in the visual axis. There are, however, general
sensation nerves underlying the conjunctiva.
- Located centrally below the
conjunctiva is the thick, transparent cornea, part of
the
outermost layer of the eyeball proper. This structure likewise lacks a
blood supply and is not innervated.The cornea is continuous with the
outer connective tissue layer of the eye known as the sclera,
the 'white' of the eye. This layer is very strong and by resisting the
fluid inflation of the eye, is important in maintaining the eye shape.
It does have vessels and sensory nerves; in addition, the orbital
muscles of the eye are inserted into the sclera.
- The innermost layer of the eye is
the retina,
which is derived embryologically from the wall
of the diencephalon of the brain. This retina is made up of many
structures, some of which are surprising: Most of the innermost layer
consists of several layers of neurons, and this is the 'visual'
retina involved in transduction of light into nervous
information and in conduction of that data to the brain. This is
the layer generally associated in people's minds with the word
'retina.'
- Toward the front of the eye,
the neural tissue of the retina has differentiated into smooth
muscle, rather than neurons and glia. A ring-shaped ciliary
body is attached to the ovoid lens
by the suspensory
ligament.
This muscle is a sphincter, and when the muscle contracts, the
tension on these ligaments is reduced, and the elastic lens
becomes more rounded; the focal length decreases...moves closer
to the eye surface. Relaxation of the muscle increases tension,
flattens the lens, and moves the focal plane farther from the
eye.
- Forward of the ciliary body,
the development of smooth muscle continues to form a two-layered disk,
the iris
diaphragm, with an opening in its center, the pupil.
The pupil restricts the incoming light to the front of the lens, where
its corrective action is best controlled and least distorted.
The muscle layers of the iris are arranged in a circle
(sphincter) or radially to change the size of the pupillary opening,
and this regulates the amount of light, the brightness, of the image.

The visual
axis is
along a line which passes straight through the apex of the cornea,
exactly through the center of the pupil, through the center of the
lens,
and finally strikes the retina. Since such a line is perpendicular to
all refractory surfaces in that pathway, there is no deflection of the
"beam." Notice the only frames of reference are in the eye structure
itself, so it doesn't matter which direction the eye is pointing.
As you
can see, any
pathway of light not on the axis is refracted
as it passes through the
eye, and at the focal point, such a beam crosses both the visual axis
and other beams (along the same circumference). The result is that it
reaches the retina on the side opposite the axis from where it entered.
This doesn't cause visual confusion, but it does cause thought
chaos
until you grow accustomed to thinking in these terms.

Thus, there are
visual
fields and
retinal
fields of vision.
- Visual fields are outside the eye.
- Retinal fields are within the eye.
- Temporal
fields are the two visual fields (one for each eye)
lateral to the visual axis.
- Medial to that axis are the two nasal
fields.
- The corresponding retinal fields are lateral
and medial.
- Designation of these sets of fields is important because of
the
frontal position of the two eyes and the degree of overlap of their
receptive fields (binocularity).
- Vision in the horizontal plane has no such special
relationship.
An upper and lower field do exist, of course, but since these do not
overlap, they are not important in visual processing.
Retinal Paths
We can teach a glass lens and plastic box to focus an image of the
correct brilliance. We call those "cameras" and some film companies
almost give them away so you will buy their film. The hard part we will
talk about here is a more advanced technology--developing that film
into (what turns out to be) a negative. This is the job of the retina,
the innermost layer of the eyeball. In the final part of this study
guide, we will send that negative along to the brain where the picture
will be printed and appreciated.
The initial processing of the image that occurs in the layers of the
retina is some of the most complex that makes up the sense of vision.
Histologists recognize 10 layers to the retina, but our approach will
be directed more to the component cells and their interrelationship.
 Rod and Cone Function
Rod and Cone Function
THESE PHOTORECEPTOR cells form a dense outer layer of the retina
(remember, the outer layer is farthest from the center of the eye
sphere).The outer segment, which is the cell region where transduction
of light actually occurs, is the outermost part of the visual retina.
- RODS
are more sensitive to light than are cones, and rods are particularly
important for night vision. The outer segment of rods is an elongate
cylinder of cytoplasm. Within this is column of discs which are
flattened membrane bubbles whose contents are technically
extracellular. These hollow wafers are produced continuously from the
basal portion of the outer segment, forming as projections of cytoplasm
that serially fuse to form the discs. These mature discs are not
connected at all to the plasma membrane.
- CONES
operate at higher light intensities and are the main receptor of
"daylight" vision, since rods saturate at very low light levels and
essentially cease to function. All color distinction is due to cone
function, based on the existence of three subtypes of cones sensitive
to three distinct light wavelengths. Rods respond only to one narrow
band of light frequency, and rod-only retinas are entirely colorblind.
Cones have a much shorter outer segment than do rods, and the
developing membranous processes that in rods fuse into the disc
series--remains unfused in the rods. This gives these cells the
appearance of bearing a kind of comb on their outer segment, but keep
in mind that in 3-D, that is really a series of plates joined at the
ciliary end.
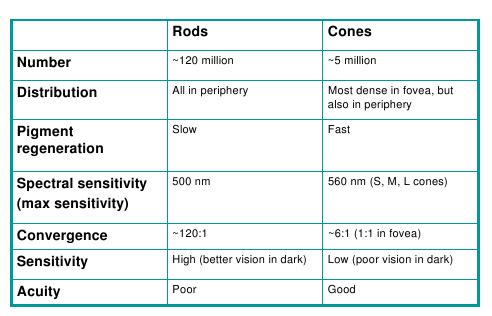
- The characteristics of the two cell types are summarized in
the following table:
 Visible Spectrum Sensitivity
Visible Spectrum Sensitivity

- Notice 3 things in the drawing above:
(1) The further away from the modal (maximum) sensitivity for each
color, the brighter the light must be before retinal receptors will
respond.
(2) Red and green overlap extensively, as do green and rod, but red has little overlap with rod wavelengths.
(3) Blue is virtually isolated from the other two colors.
Light Transduction
Both rods and cones transduce light along similar pathways, although
the pigment compounds are different in the two cell types. It is not
important at this time to go into minute chemical details of this
transduction-- rodfunctionimagethe generalities are quite sufficient.
Let's limit this discussion to rod transduction. Follow along in
the illustration below.

- Many rhodopsin molecules are mounted in the membrane of the outer
segment discs. Light of the proper frequency striking one of these is
absorbed and the molecule is excited to a higher level energetically.
- The protein opsin is embedded within the disk wall and is
attached to the light-absorbing part of its structure, retinal, a form
of vitamin A.
- When excited, the retinal dissociates from the opsin and rapidly
cascades through a series of molecular changes occupying about a
millisecond. A sequel to these changes is the activation through a
G-protein intermediate of cGMP-phosphodiesterase which reduces the
local concentration of cGMP.
- Concentration of that second messenger has been holding open the
sodium ion channels in the plasma membrane of the outer segment;
reduction of the messenger causes those gates to close, reducing the
inflow of sodium ion.
- As a result, the rod becomes more polarized--it becomes
relatively hyperpolarized, although it is really just progressing
toward its Goldman resting potential. For example, dark-adapted cones
have a transmembrane potenial of -40 mV, and exposure to light with
closure of Na+ channels plus sodium/potassium pumping raises the
potential to about -70mV. Thus, excitation of a receptor cell produces
a hyperpolarizing generator potential.
- Cones apparently behave in a similar fashion to rods in this
transduction cascade, the principal difference being that several
different opsins exist. These, in combination with the retinal molecule
absorb light at different frequencies (from that of rods), but
otherwise the response is basically the same.
Convergence: Bipolar Cells and Ganglion Cells
As we continue to examine the retina, it would be a good idea now for
you to get an appreciation of physical scale. The rods and cones are
smaller than most of the cell bodies of spinal or brain stem cells.
They are not myelinated. A proper dendrite or axon cannot be
distinguished. At their inner surface, however, they do synapse into a
column of interneurons which will process the light signal into a
sensable image. These interneurons are also quite small. The output of
their interaction with the receptor cells and with each other converges
onto ganglion cells, which are the output layer from the retina to the
brain proper. The direct throughput path is receptor to bipolar cell(s)
to ganglion. Follow the drawing below as these three cell
interrelationships are described.

As their name implies, bipolar cells have a dendritic process, a cell
body, and an axon. However, the cells are so small that these
distinctions are minor. The cells are not myelinated, and their
excitation produces an inhibitory generator potential. Synaptic input
to bipolars is from receptor cells and from a second type of
interneuron, the horizontal cells. We'll consider horizontals later.
Some of these cell interactions will seem counter-intuitive--just grit
your teeth and hang on. Rods and cones have leaky cell membranes with a
continuing inflow of sodium ion. This qualifies them as pacemaker cells
with a standing generator potential. As a result:
1. they are also leaky to calcium ions.
2. they are continuously releasing their neurotransmitter (glutmate) onto the post-synaptic membrane of the bipolars.
Bipolars are also leaky, but reception of the cone transmitter closes
those leaks--the dark adapted bipolar is inhibited. If the light
receptor is illuminated, the cone stops leaking, stops pacing...stops
releasing transmitter. The bipolar is freed from its ongoing inhibition
and depolarizes (generator), causing release of its transmitter
(glutamate) onto its ganglion cell. There are details you need to know
right now, but first you need to understand the ganglion cell.
The Ganglion cell layer is the innermost layer of the retina. Ganglion
cells have relatively large cell bodies, and from these arise long
myelinated axons that will exit the eye and make up the optic
nerve/tract synapsing in the lateral geniculate or optic tectum of
midbrain. They are leaky cells, pacemakers, but the result of this
sodium inflow is paced action potentials. Ganglion cells are inhibited
by bipolars cells which are inhibited by rods/cones which are inhibited
by light. Let's try that again:
GANGLION cells spontaneously generate action potentials that are
transmitted along the visual nerve pathway. Because each ganglion has
its own schedule independent of all the others, this input is
disorganized. Your subjective appreciation is--nothingness. The
ganglion cells can behave in this fashion because the bipolar cells
that can inhibit them are not inhibiting them because they are
inhibited by dark rod/cones.
If the light receptor cell is stimulated, it stops inhibiting the
BIPOLAR, which now begins to inhibit the ganglion cell. This imposes
order onto the discharge of the ganglion. An object has been perceived,
and as a result the ganglion communication with higher centers is
silent. You see what you don't see. Unfortunately, it's not quite that
simple. Before we go on down this maze, let's clean up three details.
- First, refer back the structure of the rods and cones. Notice
that the inner segment includes a large population of mitochondria.
These actively generate ATP to drive the cGMP transduction of light.
This role of ATP is not included in the illustration, but you should
remember this interaction from our discussion of second messenger
systems in synapses.
- Second, look at the large pathway drawing immediately above which
shows the pigmented layer of the retina outside the rod and cone layer.
The outer segments of rods and cones actually extend across the retinal
ventricle and are surrounded by a cup of pigment cells. These cups
isolate the receptors from any axis of light except straight in from
the focal point. Thus, any errant reflection or refraction which would
bounce light around in the vitreous chamber and cause false stimuli
gets filtered out. Any quanta of light that make it through the length
of the outer segment without getting absorbed, are absorbed by this
pigment layer. Many nocturnal animals develop a layer of crystals at
the floor of the pigment cup to reflect light directly back along its
axis. Thus, any light not 'seen' by receptor cells the first time get a
second shot at it. You would be aware of this structure when shining a
flashlight or headlight into such an animal's eyes--they glow. This
anatomy is called a tapetum. If you move the light slightly to the
side, the glow disappears because you are no longer along the reflected
light path--the pigment cups are absorbing that angled reflection.
- Third, a note that will be immediately obvious to you now--the
light pathway extends from the vitreous chamber, through the ganglion
cells, through the bipolar and other interneurons, and through the
length of the receptor cell BECAUSE THESE CELLS ARE TRANSPARENT.
Having just read about the pigment layer and its function, you should
understand that if polarity of the retina was reversed--if receptor
cells were the innermost layer of the retina--much incident light could
stimulate the outer segments from a variety of incoherent directions,
and image quality would be seriously fuzzy.

